
Azido erribonukleiko
| Genetika |
|---|
 |

Azido erribonukleikoa (RNA) funtsezko molekula polimerikoa da geneen kodeketan, deskodeketan, erregulazioan eta adierazpenean. RNA eta azido desoxirribonukleikoa (DNA) azido nukleikoak dira. Lipido, proteina eta karbohidratoekin batera, azido nukleikoak dira bizi mota guztientzat funtsezkoak diren lau makromolekula nagusietako bat. DNA bezala, RNA nukleotido-kate gisa eratzen da, baina DNA ez bezala, RNA bere baitan tolestutako haril bakar gisa aurkitzen da naturan, parekatutako haril bikoitz baten ordez. Organismo zelularrek RNA mezularia (mRNA) erabiltzen dute informazio genetikoa transmititzeko (G, U, A eta C letrek denotatutako guanina, urazilo, adenina eta zitosinaren base nitrogenatuak erabiliz), zeinak proteina espezifikoen sintesia zuzentzen baitu. Birus askok informazio genetikoa kodetzen dute RNA genoma bat erabiliz.
RNA molekula batzuek zeregin aktiboa dute zelulen barruan, erreakzio biologikoak katalizatzean, gene-adierazpena kontrolatzean edo zelula-seinaleei erantzunak detektatu eta komunikatzean. Prozesu aktibo horietako bat proteinen sintesia da, RNA molekulek erribosometan proteinen sintesia zuzentzen duten funtzio unibertsala. Prozesu horrek transferentziako RNA molekulak (tRNA) erabiltzen ditu aminoazidoak erribosomara eramateko; han, RNA erribosomikoak (rRNA) aminoazidoak elkartzen ditu, proteina kodetuak eratzeko.
DNArekin alderaketa
[aldatu | aldatu iturburu kodea]
DNAk bezala, biologikoki aktiboenak diren RNAek (mRNA, tRNA, rRNA, snRNAak eta beste RNA ez-kodetzaile batzuk barne) sekuentzia auto-osagarriak dituzte, eta horiei esker, RNAren zatiak bikoiztu eta parekatu egiten dira helize bikoitzak sortzeko.[3] RNA horien azterketak erakutsi du oso egituratuak direla. DNAk ez bezala, haren egiturak ez dira helize bikoitz luzeak, baizik eta elkarrekin proteinen antzeko egituretan paketatutako helize laburren bildumak.
Hala, RNAek katalisi kimikoa lor dezakete (entzimak, esaterako).[4] Adibidez, erribosomaren egitura zehazterakoan —lotura peptidiko baten eraketa katalizatzen duen RNA proteinaren konplexu bat—, ikusi zen haren gune aktiboa RNAz osatuta dagoela oso-osorik.[5]
Egitura
[aldatu | aldatu iturburu kodea]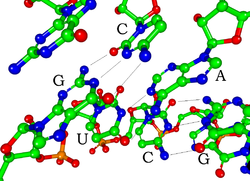
RNAren nukleotido bakoitzak azukre erribosa bat du, 1'-5' bitarteko karbonoetan. Oinarri bat finkatzen da 1' posizioan, oro har, adenina (A), zitosina (C), guanina (G) edo uraziloa (U). Adenina eta guanina purinak dira, zitosina eta uraziloa pirimidinak. Fosfato-talde bat erribosa baten 3' posizioari eta hurrengoaren 5' posizioari lotuta dago. Fosfato-taldeek karga negatibo bana dute, eta RNA molekula kargatua da (polianioia). Baseek hidrogenozko loturak eratzen dituzte zitosinaren eta guaninaren artean, adeninaren eta uraziloaren artean eta guaninaren eta uraziloaren artean.[6] Hala ere, beste elkarrekintza batzuk ere gerta daitezke, hala nola adenina-baseen talde bat, elkarri bulto batean lotzen zaizkiona, edo guanina-adenina base pare bat duen GNRA tetrabegizta.[6]

RNAren egitura-osagai garrantzitsu batek bereizten du DNAtik: talde hidroxilo bat dago azukre erribosaren 2' posizioan. Funtzio-talde horren presentziaren ondorioz, helizeak A geometria hartzen du nagusiki, nahiz eta, dinukleotido-testuinguru batean, RNAk oso gutxitan har dezakeen DNAn gehien ikusitako B forma.[7][8] A geometria ildo nagusi oso sakon eta estu batean eta azal eta zabalera txikiagoko ildo batean gertatzen da.[9] 2'-hidroxilo taldearen presentziaren beste ondorio bat da RNA molekula baten konformazio-eskualde malguetan (hau da, helize bikoitzaren eraketan sartuta ez daudenetan) kimikoki eraso dezakeela alboko fosfodiester lotura, bizkarrezurra ildaskatzeko.[10]
RNA lau basetan baino ez da transkribatzen (adenina, zitosina, guanina eta uraziloa), baina erantsitako oinarri eta azukre horiek modu askotara alda daitezke RNAak heldu ahala. Pseudouridina (Ψ), non uraziloaren eta erribosaren arteko erlazioa C–N lotura batetik C–C lotura batera aldatzen den, eta erribotimidina (T) zenbait lekutan daude (garrantzitsuenak tRNAren TΨC begiztan daude).[11] Beste base nabarmen aldatu bat hipoxantina da, nukleosina (I) izeneko adenina desaminatuko base bat.[12]
100 nukleosido natural baino gehiago daude.[13] Aldaketen egitura-aniztasun handiena ARNt-n aurki daiteke, eta pseudouridina eta 2'-O-metilribosa duten nukleosidoak dira ohikoenak.[14] RNAko aldaketa horietako askoren paper espezifikoak ez dira guztiz ulertzen. Hala ere, deigarria da ARN erribosomikoaren transkripzio osteko aldaketa asko eskualde oso funtzionaletan gertatzen direla, hala nola peptidil transferasaren zentroan eta azpiunitateen interfazean, eta horrek esan nahi du garrantzitsuak direla funtzio normalerako.[15][16]
Sekzio bakarreko RNA molekulen forma funtzionalak, proteinek bezala, sarritan egitura tertziario espezifikoa eskatzen du. Egitura horretarako aldamioa molekularen barruan hidrogeno-loturak diren bigarren mailako egitura-elementuek osatzen dute. Horren ondorioz, egitura sekundariotik ezagutzen ahal diren "eremu" batzuk sortzen dira, hala nola urkila begiztak, fardelak eta barne begiztak.[17] Bigarren mailako edozein egituratarako RNA bat sortzeko, bi edo hiru oinarri ez lirateke aski izango, baina lau oinarri nahikoak dira.[18] Litekeena da naturak lau baseko alfabetoa "aukeratu" izana: lau baino gutxiagok ez du egitura guztiak sortzeko aukerarik ematen, eta lau oinarri baino gehiago ez dira beharrezkoak. RNA kargatzen denez, ioi metalikoak behar dira, hala nola Mg2+, bigarren eta hirugarren mailako egitura asko egonkortzeko.[19]
RNAren enantiomero naturala D-ARN da, D-erribonukleotidoz osatua. [20]
Sintesia
[aldatu | aldatu iturburu kodea]RNAren sintesia entzima batek katalizatu ohi du —RNA polimerasak—, DNA txantiloi gisa erabiliz, transkripzio deritzon prozesu gisa. Transkripzioaren hasiera entzima DNAko sekuentzia sustatzaile batera elkartzean hasten da (normalean, gene batetik "korrontean gora" egoten da). Entzimaren helikasa jarduerak DNAren helize bikoitza eragiten du. Ondoren, txantiloiaren katean zehar, entzimak 3’-tik 5’-ra egiten du aurrera, RNA molekula osagarri bat sintetizatuz, 5’-tik 3’-ra doan norabidean gertatzen den elongazioarekin. DNAren sekuentziak RNAren sintesia non amaituko den ere adierazten du.[21]
RNA-mendeko RNA polimerasen kopuru batek ere RNA erabiltzen du RNA kate berri baten sintesirako. Adibidez, zenbait RNA birusek (poliobirusak, esaterako) entzima mota hori erabiltzen dute material genetikoa erreplikatzeko.[22] Gainera, RNAren mendeko RNAren polimerasa RNAk organismo askotan duen interferentzia-bidearen parte da.[23]
Aurkikuntza
[aldatu | aldatu iturburu kodea]XIX. mendearen bukaeran, Kossler ikerlaria konturatu zen zenbait zelularen nukleotik isolatutako nukleina deritzen substantzien osagaietan base nitrogenatu desberdinak ageri zirela. Timoko zeluletan, adibidez, timina agertzen zen; legamietakoetan, ordea, uraziloa. Beraz, lehenengo kasuan, isolatutakoa DNA-molekula izango zen; bigarrenean, aldiz, RNA.
XX. mendearen erdialdean, Severo Ochoak, RNA-kateak eratzeko nukleotidoak lotzen dituen RNA-polimerasa entzima aurkitu ondoren, laborategian RNA-molekula sintetizatzea lortu zuen. Azido erribonukleikoa bere osagaietatik abiatuta eratzeko bidea aurkitzeagatik Nobel saria eman zioten 1959. urtean. Orain dela gutxi ere, 2006. urtean, RNA-interferentziarekin lanean zebiltzan ikertzaileak saritu zituzten geneen espresioen isiltze-prozesua azaltzearren[24].
RNA motak
[aldatu | aldatu iturburu kodea]RNA mezularia (RNAm)
[aldatu | aldatu iturburu kodea]Bere izenak (mezularia) agerian jartzen du bera dela DNAn dagoen informazio genetikoa erribosometara garraiatzen duena, bertan proteinak sortzeko.
Hari bakarreko kate lineala da. Nukleoan sortzen da transkripzio izeneko prozesuan. Prozesu horretan RNA polimerasa entzimak DNAren bi kateak banandu eta kate bat moldetzat hartuz erribonukleotidoak gehitzen ditu, RNAm-aren kate berria agertuz. Sortzen ari den RNAaren kate berrian DNAren moldearekiko osagarriak diren base nitrogenatuak eta nukleotidoak jartzen ditu RNA polimerasak (adeninaren aurrean urazilo jarriz, eta guaninaren aurrean zitosina).
RNA mezulariak proteinen sintesirako kode genetikoa eramaten duenez, RNAm-ari kodifikatzaile deritzo. Beste RNA motak (RNAr, RNAt), aldiz, ez dira kodifikatzaileak.
Itzulpen prozesuan RNAm-aren nukleotido hirukoteek (kodoiek) aminoazido bat kodetuko dute.
RNA erribosomikoa (RNAr))
[aldatu | aldatu iturburu kodea]Hainbat proteinekin lotzerakoan erribosomen egitura osatzen du. Erribosomen masa guztiaren %65a RNAr-ari dagokio.
Erribosomak proteinen ekoizpenaren zentruak dira. Bi azpiunitatez osatuta daude (handia eta txikia), eta ezberdinak dira zelula prokariotoetan eta zelula eukariotoetan. Prokariotoen erribosometan 3 RNAr ezberdin daude, eta eukariotoetan, 4.
RNA mota honek ezaugarri katalitikoak ditu itzulpen prozesuan; beraz, erribozimen antzera jarduten du.
RNA transferentziazkoa (RNAt)
[aldatu | aldatu iturburu kodea]
Hirusta-hosto baten morfologia du. Aurrez aurre eta hidrogeno zubien bidez lotuta dauden RNAren kate bikoitzak ditu, lau beso osatuz (ikus irudia). Hiru bukle ditu beso horien muturretan, bertan ez dagoelako baseen arteko loturarik.
Goiko besoan kate bat bestea baino luzeagoa da. Hain zuzen, kate luzeago horretan ACC baseak daude. Toki horri atxikitzen zaio RNAt guztietan dagoen aminoazidoa (3' muturrean).
Beheko besoan, aldiz, antikodoi izeneko tokian 3 base nitrogenatu dauzka. Goiko besoan dagoen aminoazidoa antikodoian dauden 3 baseen arabera izango da. Hots, antikodoi bakoitzari aminoazido bat dagokio.
Era berean, antikodoian dauden 3 base horiek RNAm-aren 3 base osagarriekin (kodoian daudenekin) lotuko dira erribosometan, itzulpen prozesua burutzen denean. Nabaria da, beraz, funtsezko zeregina daukala RNAt-k proteinen sintesian, bera baita RNAm-an dagoen kode genetikoari aminoazido espezifiko bat jartzen diona.
RNAt-ren zenbait eremuk egitura primarioa ez ezik, egitura sekundarioa ere badute (aipatu den bezala baseen arteko lotura kimikoak ezartzen baitira eta helize bikoitza sortzen baitute).
Eboluzioa
[aldatu | aldatu iturburu kodea]Zenbait ikertzaileren arabera, bizia sortu zen ingurunean RNA-molekulak egokiak izango ziren informazio genetikoaren gordailu eta garraiatzaile gisa; izan ere, funtzio horiek betetzeaz gain, aktibitate katalizatzailea ere badute. Azido erribonukleikoaren ezegonkortasuna dela eta, zelulek bestelako molekula egonkorragoak eratuko zituzten hain garrantzi handia duen informazioa gordetzeko, azido desoxirribonukleikoa, hain zuzen ere. Katalisiari dagokionez, askotariko egiturak dituzten molekulak eratuko ziren, hau da, proteinak. Egitura horiei esker, funtzio gehiago betetzeko gai izango ziren. Egungo zeluletan, hiru molekula-mota horiek, DNAk, proteinek eta RNAk, betetzen dituzte egokien dagozkien funtzio bereiziak, hots, informazio-gordailu, katalizatzaile eta bien arteko bitartekari izatea[24].
Ikus, gainera
[aldatu | aldatu iturburu kodea]Erreferentziak
[aldatu | aldatu iturburu kodea]- ↑ RNA: The Versatile Molecule. University of Utah.
- ↑ Nucleotides and Nucleic Acids. University of California, Los Angeles.
- ↑ «How RNA folds» Journal of Molecular Biology 293 (2): 271–81. October 1999 doi:. PMID 10550208..
- ↑ «RNA secondary structure: physical and computational aspects» Quarterly Reviews of Biophysics 33 (3): 199–253. August 2000 doi:. PMID 11191843..
- ↑ «The structural basis of ribosome activity in peptide bond synthesis» Science 289 (5481): 920–30. August 2000 doi:. PMID 10937990. Bibcode: 2000Sci...289..920N..
- ↑ a b «Diversity of base-pair conformations and their occurrence in rRNA structure and RNA structural motifs» Journal of Molecular Biology 344 (5): 1225–49. December 2004 doi:. PMID 15561141..
- ↑ «The DNA strand in DNA.RNA hybrid duplexes is neither B-form nor A-form in solution» Biochemistry 32 (16): 4207–15. April 1993 doi:. PMID 7682844..
- ↑ «RNA approaches the B-form in stacked single strand dinucleotide contexts» Biopolymers 105 (2): 65–82. February 2016 doi:. PMID 26443416..
- ↑ «RNA bulges as architectural and recognition motifs» Structure 8 (3): R47–54. March 2000 doi:. PMID 10745015..
- ↑ «The mechanism of the metal ion promoted cleavage of RNA phosphodiester bonds involves a general acid catalysis by the metal aquo ion on the departure of the leaving group» Journal of the Chemical Society, Perkin Transactions 2 (8): 1619–26. 1999 doi:..
- ↑ «Identification of critical elements in the tRNA acceptor stem and T(Psi)C loop necessary for human immunodeficiency virus type 1 infectivity» Journal of Virology 75 (10): 4902–6. May 2001 doi:. OCLC .114245 PMID 11312362..
- ↑ «Inosine biosynthesis in transfer RNA by an enzymatic insertion of hypoxanthine» The Journal of Biological Chemistry 259 (4): 2407–10. February 1984 doi:. PMID 6365911..
- ↑ «The RNA Modification Database, RNAMDB: 2011 update» Nucleic Acids Research 39 (Database issue): D195-201. January 2011 doi:. OCLC .3013656 PMID 21071406..
- ↑ «Small nucleolar RNA-guided post-transcriptional modification of cellular RNAs» The EMBO Journal 20 (14): 3617–22. July 2001 doi:. OCLC .125535 PMID 11447102..
- ↑ «The Peptidyl Transferase Center: a Window to the Past.» Microbiol Mol Biol Rev 85 (4): e0010421. November 2021 doi:. OCLC .8579967 PMID 34756086..
- ↑ «Ribosome structure and activity are altered in cells lacking snoRNPs that form pseudouridines in the peptidyl transferase center» Molecular Cell 11 (2): 425–35. February 2003 doi:. PMID 12620230..
- ↑ «Incorporating chemical modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure» Proceedings of the National Academy of Sciences of the United States of America 101 (19): 7287–92. May 2004 doi:. OCLC .409911 PMID 15123812. Bibcode: 2004PNAS..101.7287M..
- ↑ «RNA secondary structure design» Physical Review E 75 (2): 021920. February 2007 doi:. PMID 17358380. Bibcode: 2007PhRvE..75b1920B..
- ↑ «Salt dependence of nucleic acid hairpin stability» Biophysical Journal 95 (2): 738–52. July 2008 doi:. OCLC .2440479 PMID 18424500. Bibcode: 2008BpJ....95..738T..
- ↑ «Turning mirror-image oligonucleotides into drugs: the evolution of Spiegelmer(®) therapeutics» Drug Discovery Today 20 (1): 147–55. January 2015 doi:. PMID 25236655..
- ↑ «Transcription termination and anti-termination in E. coli» Genes to Cells 7 (8): 755–68. August 2002 doi:. PMID 12167155..
- ↑ «Structure of the RNA-dependent RNA polymerase of poliovirus» Structure 5 (8): 1109–22. August 1997 doi:. PMID 9309225..
- ↑ «RNA-dependent RNA polymerases, viruses, and RNA silencing» Science 296 (5571): 1270–73. May 2002 doi:. PMID 12016304. Bibcode: 2002Sci...296.1270A..
- ↑ a b «ZT Hiztegi Berria» zthiztegia.elhuyar.eus (Noiz kontsultatua: 2018-12-17).
Kanpo estekak
[aldatu | aldatu iturburu kodea]| Autoritate kontrola |
|
|---|